生命物理化学研究室の研究概要
1、研究テーマの全般的な説明
 ポストゲノム時代に特に注目されている生命科学研究において、タンパク質が生体内で相手をどのようにして認識し識別するか明らかにすることを目的としています。生体内において遺伝情報がアミノ酸に翻訳され、アミノ酸の並びであるタンパク質が立体構造を形作り、個々の機能を発現しています。ポストゲノム研究において世界的にタンパク質の立体構造解析が進められていますが、我々は特にタンパク質の溶液中における揺らぎの寄与に着目し、このタンパク質の動的構造と機能との相関を明らかにしていきたいと考えています。対象は抗体、免疫系シグナル伝達関連タンパク質、各種酵素などで、個々のタンパク質が相手をどのように認識するか、結合する際の構造変化を含む動的構造要因は何か、などを解明し、生体システムを理解していきたいと、分子レベルからのアプローチで生命科学研究を行っています。さらにこれら基礎研究の成果が、結果として医薬品や研究手法の開発など、応用面でも貢献できればと考えています。
ポストゲノム時代に特に注目されている生命科学研究において、タンパク質が生体内で相手をどのようにして認識し識別するか明らかにすることを目的としています。生体内において遺伝情報がアミノ酸に翻訳され、アミノ酸の並びであるタンパク質が立体構造を形作り、個々の機能を発現しています。ポストゲノム研究において世界的にタンパク質の立体構造解析が進められていますが、我々は特にタンパク質の溶液中における揺らぎの寄与に着目し、このタンパク質の動的構造と機能との相関を明らかにしていきたいと考えています。対象は抗体、免疫系シグナル伝達関連タンパク質、各種酵素などで、個々のタンパク質が相手をどのように認識するか、結合する際の構造変化を含む動的構造要因は何か、などを解明し、生体システムを理解していきたいと、分子レベルからのアプローチで生命科学研究を行っています。さらにこれら基礎研究の成果が、結果として医薬品や研究手法の開発など、応用面でも貢献できればと考えています。
2、個々の研究テーマの簡単な説明
2-1、抗体と抗原
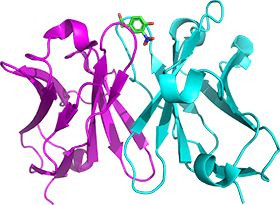 抗体は同様の立体構造を持ちながら、様々な抗原を認識し、結合しうるタンパク質で、最近では抗体医薬など応用面でも広く利用されています。この多様な抗原認識機構解明をはじめ、安定性の向上、凝集性の抑制など、基礎研究として解決すべく課題も多く、我々は抗原結合部位を含む 抗体の可変領域を1本鎖Fv(scFv)抗体として調製し、抗体の低分子化、アミノ酸置換体の創製といったscFvの利点を活かし、様々な物理化学的解析手法を通じて、抗原結合に伴う構造変化の寄与の解明などに取り組んでいます。
抗体は同様の立体構造を持ちながら、様々な抗原を認識し、結合しうるタンパク質で、最近では抗体医薬など応用面でも広く利用されています。この多様な抗原認識機構解明をはじめ、安定性の向上、凝集性の抑制など、基礎研究として解決すべく課題も多く、我々は抗原結合部位を含む 抗体の可変領域を1本鎖Fv(scFv)抗体として調製し、抗体の低分子化、アミノ酸置換体の創製といったscFvの利点を活かし、様々な物理化学的解析手法を通じて、抗原結合に伴う構造変化の寄与の解明などに取り組んでいます。
2-2、T細胞補助刺激受容体CD28とシグナル伝達アダプター分子
 T細胞活性化 には2つの独立したシグナル、1)主要組織適合抗原複合体(MHC)と結合するT細胞受容体(TCR)経由、2)CD28などの補助シグナル分子経由、が必要です。我々は主に2)について、CD28、ICOS、CTLA-4など各分子の細胞内領域にあるリン酸化チロシンを含む領域と、同領域に結合する各種シグナル伝達タンパク質のSH2との分子間相互作用に着目し、様々な構造機能解析を通じて、シグナル伝達機構を解明したいと考えています。
T細胞活性化 には2つの独立したシグナル、1)主要組織適合抗原複合体(MHC)と結合するT細胞受容体(TCR)経由、2)CD28などの補助シグナル分子経由、が必要です。我々は主に2)について、CD28、ICOS、CTLA-4など各分子の細胞内領域にあるリン酸化チロシンを含む領域と、同領域に結合する各種シグナル伝達タンパク質のSH2との分子間相互作用に着目し、様々な構造機能解析を通じて、シグナル伝達機構を解明したいと考えています。
なお本研究の結晶構造解析は、JAXAの高品質タンパク質結晶生成実験としても進めています。
2-3、エンド-1,3-β-グルカナーゼ
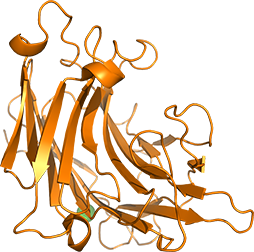 N末端側に触媒ドメインを、C末端側に糖結合モジュールを有するCellulosimicrobium cellulans由来のエンド-1,3-β-グルカナーゼについて、各ドメインの構造機能相関解析を行っています。各ドメインが如何に糖を認識し、機能するかを解明することで、触媒活性や糖結合能の高機能化を目指します。
N末端側に触媒ドメインを、C末端側に糖結合モジュールを有するCellulosimicrobium cellulans由来のエンド-1,3-β-グルカナーゼについて、各ドメインの構造機能相関解析を行っています。各ドメインが如何に糖を認識し、機能するかを解明することで、触媒活性や糖結合能の高機能化を目指します。
2-4、クチナーゼ様酵素Cut190
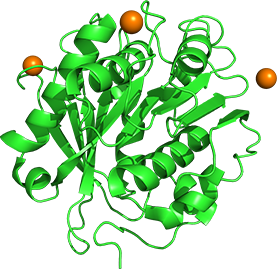 Saccharomonospora viridis AHK190由来のクチナーゼ様酵素Cut190は、Ca2+存在下で、安定化、および活性化し、polyethylene terephthalate (PET) を分解します。
Saccharomonospora viridis AHK190由来のクチナーゼ様酵素Cut190は、Ca2+存在下で、安定化、および活性化し、polyethylene terephthalate (PET) を分解します。
Ca2+結合に伴うCut190の立体構造変化は、これまでの複数の結晶構造でも明らかで、今後はさらにCa2+結合に伴う動的構造変化を解明し、安定性や機能との相関を明らかにして、PET分解酵素の高機能化にも繋げたいと考えています。
2-5、リボヌクレアーゼHI
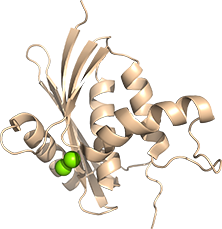 リボヌクレアーゼHIは、RNA/DNA hybridのRNA鎖を加水分解する酵素です。活性に二価金属イオンを要し、Mg2+存在下で最も活性が高いことや、Mn2+、Zn2+、Co2+存在下でも活性があること、Ca2+存在下では活性がみられないことが報告されています。この金属イオンによる活性制御は、既存の結晶構造解析では見えていない動的挙動や、付随する酵素の動的構造変化が関係すると考えられ、酵素の繊細かつ動的な構造機能相関の解明を目指しています。
リボヌクレアーゼHIは、RNA/DNA hybridのRNA鎖を加水分解する酵素です。活性に二価金属イオンを要し、Mg2+存在下で最も活性が高いことや、Mn2+、Zn2+、Co2+存在下でも活性があること、Ca2+存在下では活性がみられないことが報告されています。この金属イオンによる活性制御は、既存の結晶構造解析では見えていない動的挙動や、付随する酵素の動的構造変化が関係すると考えられ、酵素の繊細かつ動的な構造機能相関の解明を目指しています。
2-6、DNA結合タンパク質
転写因子c-MybのDNA結合ドメインは、R1, R2, R3と呼ばれる3つのリピートから成り、このうちR2R3が、特異的DNA結合に必須であることが知られています。これまでの一連の研究で、このR2R3は、天然変性状態に近い構造柔軟性を有しており、この「柔らかさ」が機能発現に重要であることを明らかにしました。今後はさらに「柔らかさ」に着目した構造機能相関解明を進めたいと考えています。
2-7、ヘリックスバンドル形成タンパク質
ヒスチジンの金属結合性を利用し、Zn2+などの金属イオン存在下で、ランダムコイル状態から、ヘリックスバンドル構造に変化するタンパク質を設計、創製しています。これらタンパク質を用いて、構造変化に伴う運動性の解析を1分子レベルでも行い、アンサンブル量となる他の物理化学的解析結果とも相関付け、タンパク質のフォールディングや分子間相互作用の基礎情報を得たいと考えています。
3、研究の進め方の概要
3-1、タンパク質の調製
まずは実験に使う試料を準備します。タンパク質は主に大腸菌の大量発現系により産生させますが、そのために必要な遺伝子をPCR法で増幅したり、制限酵素でDNAを切断し目的の遺伝子を組み込んだりして、プラスミドDNAを調製します。次にプラスミドDNAを導入した大腸菌を培養することで得られる目的タンパク質を、種々のクロマトグラフィーを用いることで純度の高いものに精製します。
また比較的小さなタンパク質は化学合成によっても調製します。
3-2、タンパク質の物性解析
従来手法によるタンパク質の安定性解析や酵素反応解析の他にも次のような機器を用いています。
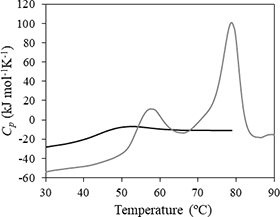
1) 示差走査熱量計
高感度でタンパク質など生体高分子の安定性や変性に伴う熱力学量を求めることができます。
2) マススペクトロメトリー
タンパク質など生体高分子の分子量を高感度で決定でき、さらに結合解析にも適用できます。
3-3、タンパク質の分子間相互作用解析
電気泳動法やゲルろ過クロマトグラフィー以外にも次のような機器を用いています。
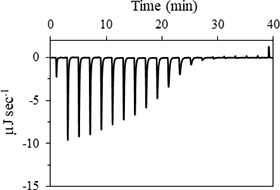
1) 等温滴定熱量計
生体高分子-リガンド相互作用における微少な吸発熱を超高感度測定し、結合に伴う熱力学量や結合比を求めることができます。
2) 表面プラズモン共鳴バイオセンサー
比較的微量のタンパク質試料で、結合の強さや結合速度を求めることができます。
3) ストップトフロー法
比較的速い反応(ミリ秒またはそれ以下)を速度論的に解析することができます。
3-4、タンパク質の立体構造解析
主に次のような機器を用いて行います。
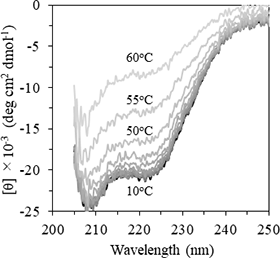
1) CD(円偏光二色性)スペクトル解析
比較的微量のタンパク質試料で二次構造を解析することができます。
2) X線結晶構造解析
タンパク質を結晶化し、その立体構造を数Åの分解能で同定します。
3) NMR(核磁気共鳴)解析
溶液中でのタンパク質など生体高分子の立体構造解析が可能です。
