※rps14遺伝子の場合※ (Kubo et al. 1999)
・rps11遺伝子の場合と同様に、イネのrps14遺伝子も進化的に比較的新しくミトコンドリアから核へ転移したと推定された。
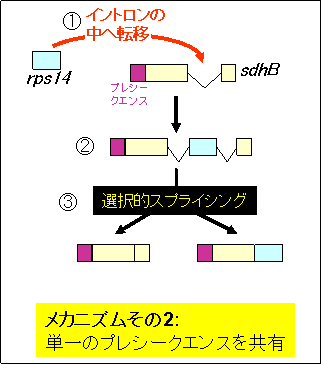
① 新たに転移したrps14遺伝子は、核で既に機能を獲得している別のミトコンドリア遺伝子(sdhB)の内部に転移した。
② これによって、二つの遺伝子はプレシークエンスを共有することになった。
③ それぞれの遺伝子は、選択的スプライシング(イントロンのスプライシングパターンの変化)によってmRNAが生成される。
※rps11遺伝子の場合※ (Kadowaki et al. 1996)
・イネのrps11遺伝子は進化的に見て比較的新しい時期にミトコンドリアから核へ転移したと推定された。
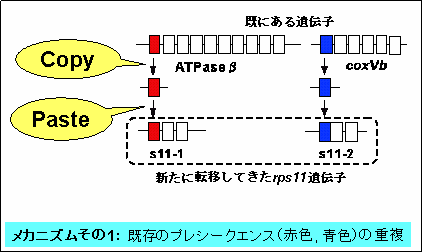
・新たに転移したrps11遺伝子は、核で既に機能を獲得している別のミトコンドリアの遺伝子(atpBとcoxVb)からプレシークエンスがコピーされたと考えられる。これは、プレシークエンスの進化的な由来と獲得のメカニズムが判明した初めての例である。
ミトコンドリアの遺伝子転移
無断転載を禁ず
以上の2例から、新たに核へ転移したミトコンドリアの遺伝子は、既に核で機能を獲得した遺伝子を利用して効率的に進化してきたことが明らかとなった(身近な例で例えると、rps11の場合が“盗作”(例えは悪いですが)、rps14の場合が“家の同居”に相当すると思います)。
<ミトコンドリアから核へ転移した遺伝子の進化>
ミトコンドリアの機能にとって必要な遺伝子の大部分は核に存在し、細胞質での翻訳後にミトコンドリアへ輸送される。このような核の遺伝子の多くは「かつてはミトコンドリアゲノムにあったが、進化の過程で核ゲノムへ転移したもの」であると考えられている。
ミトコンドリアから核へ転移した遺伝子は、長い進化の時間の中で、核の中で機能するために必要な機構を獲得していったのであろうが、その過程はほとんど謎のままである。
最近の研究から、既に大多数の遺伝子を転移させた動物とは異なり、植物では核への遺伝子転移が現在も進行中であることが報告されている。
このような植物ミトコンドリアの特徴を利用して、核へ転移して比較的新しい遺伝子を単離した。それらの解析の結果、転移した遺伝子の活性化の過程が明らかとなった。特に、ミトコンドリアへのタンパク質の輸送には、“プレシークエンス”と呼ばれる輸送シグナルが必要である(前頁の図4参照)。下記の解析ではプレシークエンスの由来と獲得のメカニズムが初めて明らかにされた。
※rpl6遺伝子の場合※ (Kubo et al. 2008)
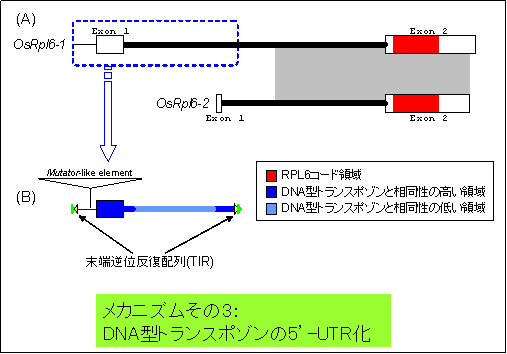
・核に転移したイネrpl6遺伝子は、2コピー存在する(上図の(A))。
・それらのうちの一方のコピー(OsRpl6-1)の5'-非翻訳領域(UTR)は、DNA型トランスポゾンに特徴的な構造を持つ(B)。