このページは 旧所属(北海道大学低温科学研究所 寒冷域植物生理生態分野)に
所属していたとき作成したものに修正を加えたものです。。
隅田明洋のページ:
 Photo taken in 2012
Photo taken in 2012
隅田 明洋
専門 森林生態学
profile 生まれと育ち 長崎市
1978 長崎西高等学校 卒(陸上部)
1983 名古屋大学 卒(農学部)
1993 学位取得(名古屋大学)
1993 - 1996 科学技術特別研究員(PD) 森林総合研究所関西支所
1997 - 1999 岐阜大学 助手(農学部)
1999 - 2020 北海道大学 准教授(低温科学研究所)
2020 - 現在 京都府立大学大学院 教授(生命環境科学研究科)
研究の内容 (たまに更新)
これまでの研究トピックス:
研究トピック(1) :風と森林の高さについての話。
研究トピック(2)-1 :樹木の形の法則と葉の量との関係についての話。
研究トピック(2)-2 :森林の葉量垂直分布を地上からレーザー距離計で測る話。
研究トピック(3) 幹の形ができる過程を明らかにした論文が
Tree Physiology 誌に掲載されました(下記)) →Open Access
研究トピック(4) 常緑針葉樹林の葉面積指数の年変動と気象要因との関係についての論文が
Scientific Reportsに掲載されました →Open Access
Publication (English & Japanese)
生き物の形には自然の中で生存するための戦略が隠れています。
森林の中の樹木の形(樹形)は、樹木の生存や森林の維持に対してどのようなかかわりがあるのでしょうか?
それを明らかにするために、
- 樹形(木の幹の太さや長さ,枝の構造,枝葉の広がりなどの三次元的構造)が、木の種類や成長の段階、
木をとりまく環境などによってどう変わるか? - 樹形はどのように発達あるいは維持されているのか?
- 樹形の発達の仕方が木々の間の生存競争とどのようにかかわっているのか?
- 樹形の発達の仕方は,森林全体の構造やその時間変化(森林動態)のパターンとどのように
かかわっているのか? あるいはかかわっていないのか?
・・・などを調べ、森林の生態について理解を深めようとしています。
森林のなかの個体間の幹の形の形成過程の違いと樹冠との関係
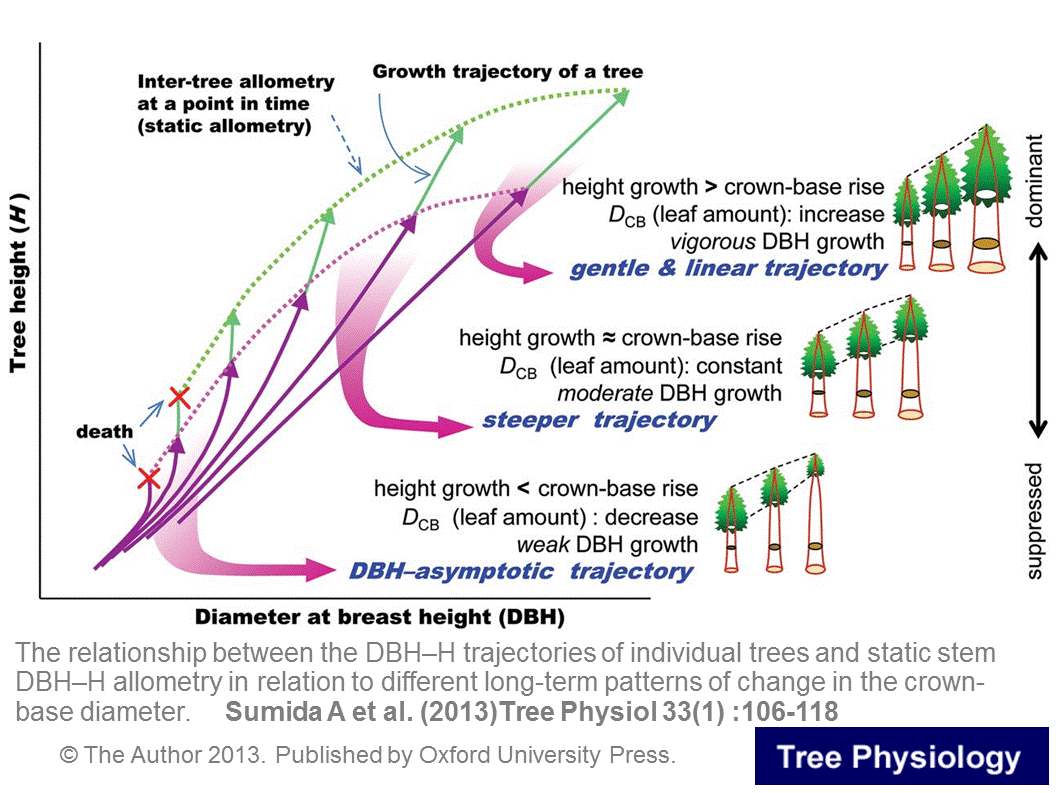
上の図は、幹の太さ(DBH:高さ1.3mの位置の幹の太さ)と樹高(H)との関係が時間とともにどのように変化していくか、を明らかにした論文(Sumida et al. 2013)の内容です。森林内には小さな個体や大きな個体が生存していますが、それらの樹冠の大きさの樹齢にともなう移り変わりのしかたが幹の形に反映されることを明らかにしました。さらに、森林全体の葉の量と気象との関係についての研究や、樹冠の時間変化を枝レベルで追跡する研究などを行っています。
興味のはじまりはちょっとした疑問から
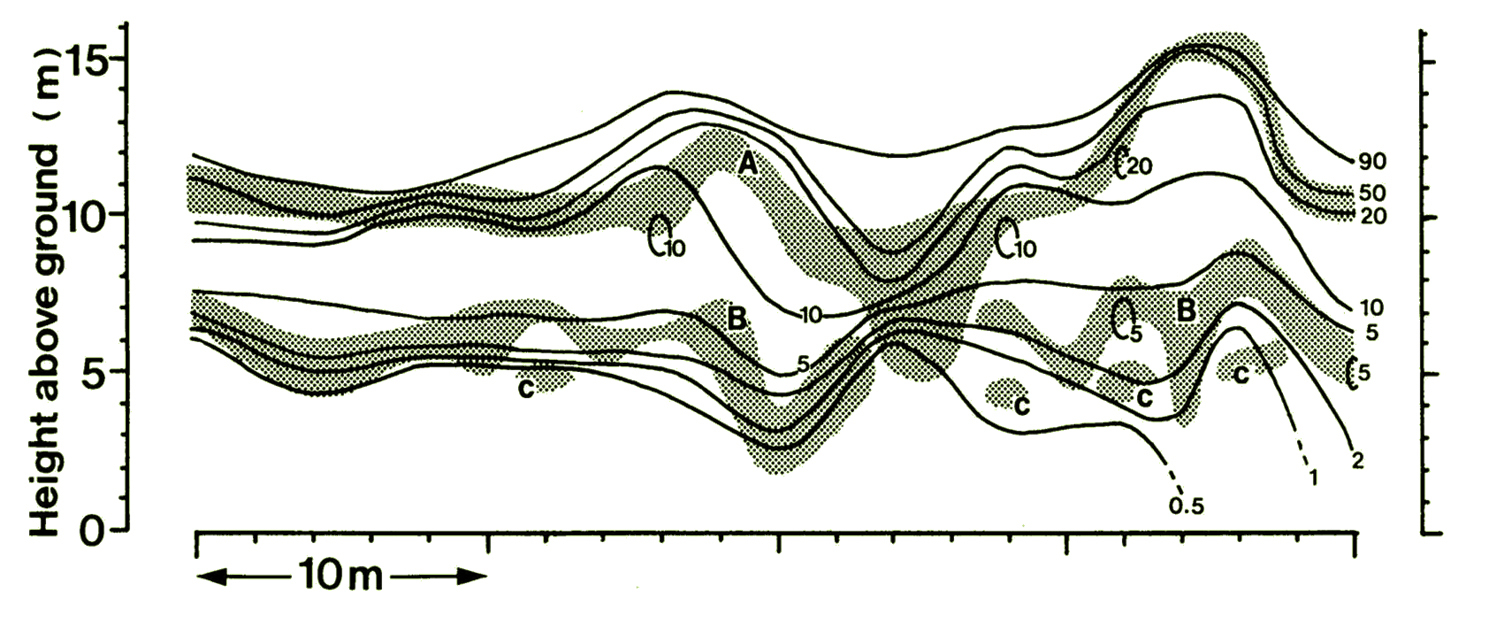
私が大学院生の頃、よく木登りをしました(研究のため)。森林の林冠(表層の、葉がこんもり茂っているところ)まで登ると、たくさんの樹木の葉があつまって層状になり、下側の層はでこぼこな緑のじゅうたんのようでした(下図)。
広葉樹林の林冠構造の断面図。20cmの厚さで森林をスライスしたように表現した。グレーの部分は葉が詰まった空間(葉層)。相対的な明るさ(%)の等値線は、葉層のある位置で間隔が狭くなっている。つまり、光が葉によって吸収されている。葉がないところでも明るさが減衰しているのは、この図の外側にある葉によって光が吸収されているため。オリジナルデータ; Sumida (1993), Sumida (1995)).
上のような葉群の層状構造は、層構造を保ちながら時間とともに上へ上へと発達していきます。このことは、樹木がどのように大きくなるのかを知っていると、ちょっと不思議です。
枝のはじまりから林冠の発達を考える
樹木は、当年性シュート(毎年伸びる、葉がついた小枝)を親枝から分岐させながら成長します。
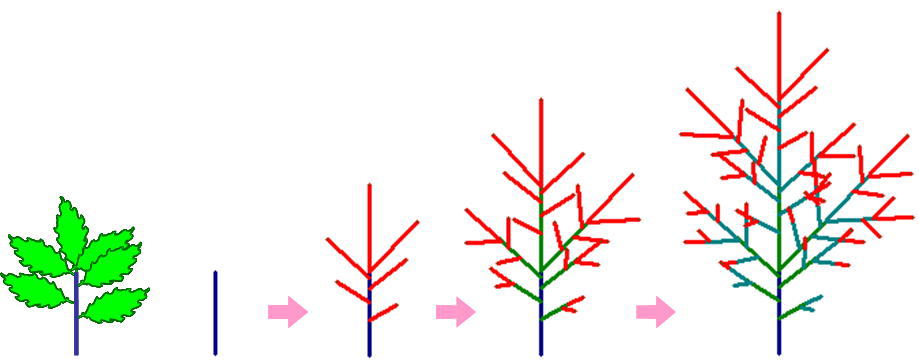
一本の当年性シュート(左)が樹冠の中である年に生まれると、その枝は翌年こんどは親枝になり、複数の娘枝(赤い線で描いた当年性シュート:葉は描いてない)を生みます。さらに、それらの娘枝はそれぞれ次の年に親枝となって当年性シュートを生む・・・を繰り返します。

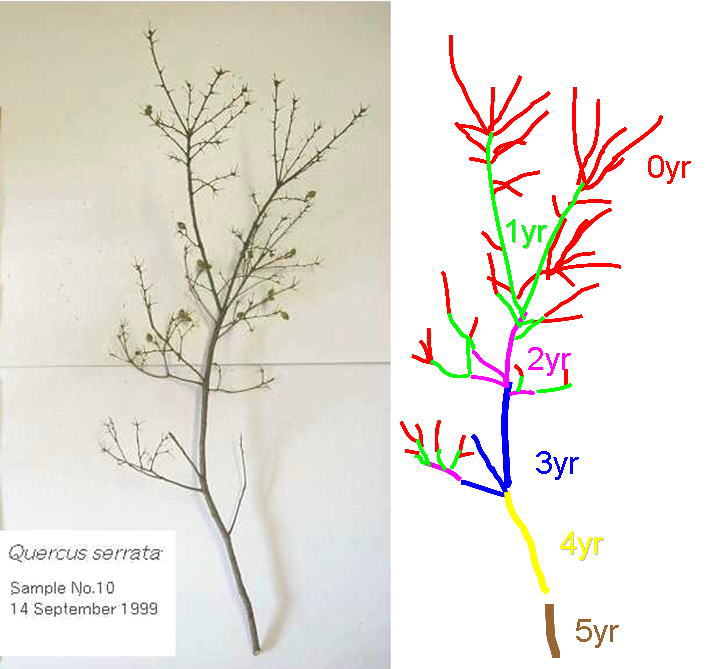

一本の親枝が複数の娘枝を生む、ということを繰り返すと枝の数はどうなるでしょうか?樹冠のいちばん上のほうから枝を採取してきて、葉を取り除き(上の左の写真)、枝の各部分の年齢ごとに本数を数えてやります(上の右図のように)。そして、縦軸に年齢別のシュートの本数(y)、横軸に枝の年齢(x)をとったグラフを描いてみました。それが下の図です。
縦軸を対数目盛り(数値の増加割合が一定となるようにとった目盛り)にとってやると、きれいな直線関係が得られます(下図:データは上の写真とは別の枝のもの)。
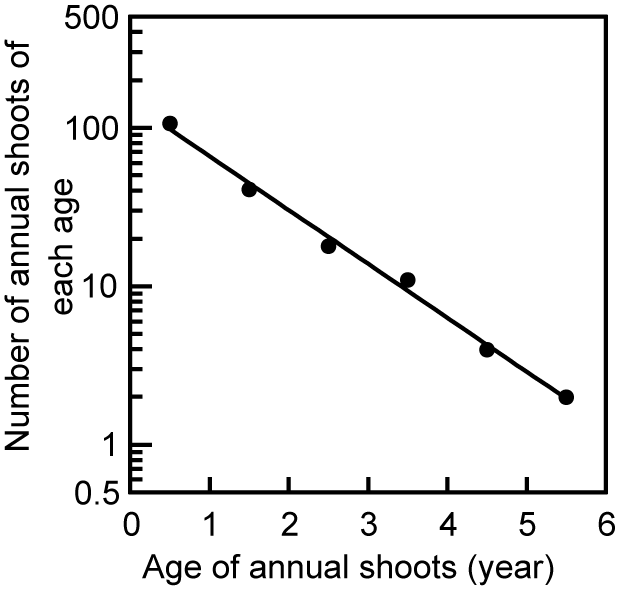
Figure 1 in Sumida & Takai (2003)
この直線関係は、xとyとの間に指数関数的関係(式でかくと y = a×e-b x ; eは自然対数の底を表す定数))が成立していることを示しています。また、この直線の傾き(式の「b」の値に相当)は、枝の年齢が1年若くなるごとに当年性シュートの本数が2~3倍に増えることを示しています。
しかし、森林の林冠が閉鎖(下の写真のように、樹冠どうしが隙間無く埋まったような状態にあること)してしまうと、葉がついた当年性シュートの本数を指数関数的に増やせる空間はありません。

実際、森林の林冠が閉鎖した後は、森林全体の葉の量がほぼ一定になる(それ以上増えない状態になる)ことはよく知られています。ではいったい、枝分かれして小枝の本数を増やし続ける性質をもつ樹木と、森林の葉量が一定となる性質とはどのように両立しているのでしょうか?
みかけは指数関数的発達、じつは定常状態
答えのひとつは意外と簡単かもしれません。一本の当年性シュートが親枝となって枝が発達し続けるとき、その親枝のいちばん先端のシュート(下の図で「Apical」と書いたシュート)由来の部分だけが成長を続け、おなじ親枝のそれ以外のシュート(斜め向きに生えている「側枝」)由来の枝はだんだん発達の度合いが悪くなり、最後は芽を付けなくなってしまう(つまり翌年死んでしまう)、という現象がおこっていることがわかりました。
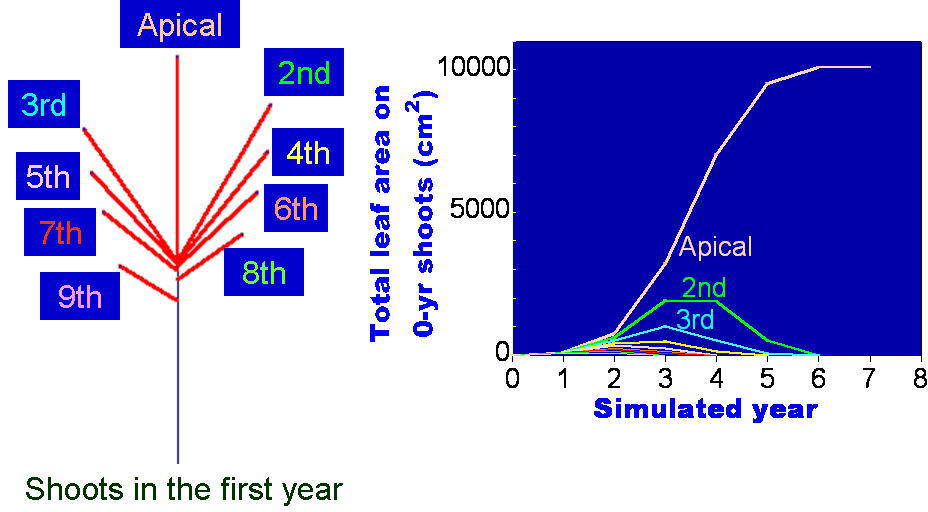
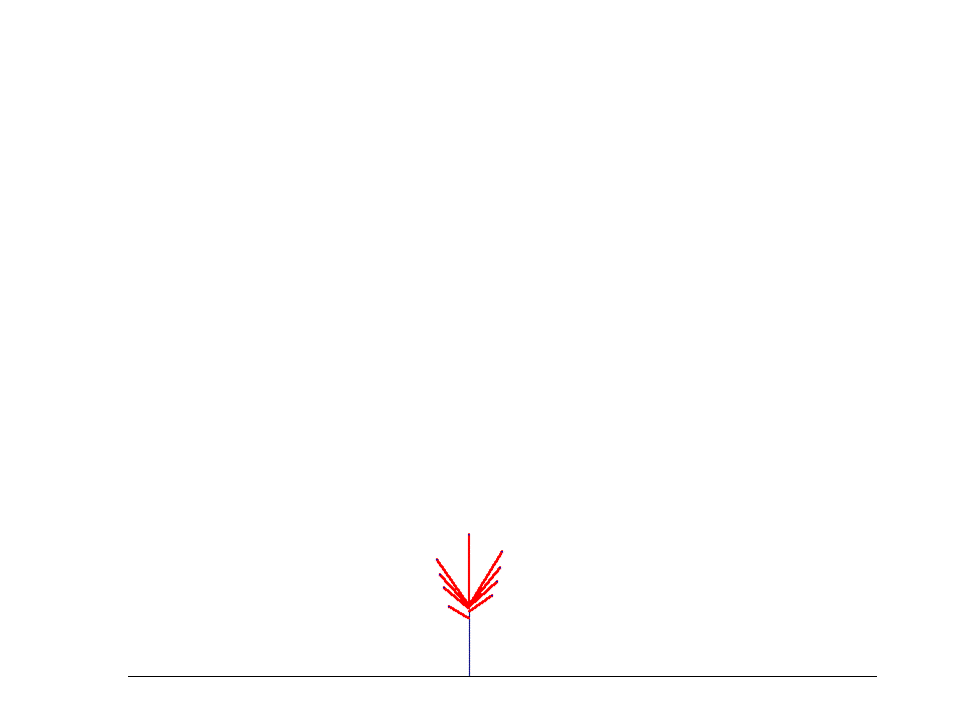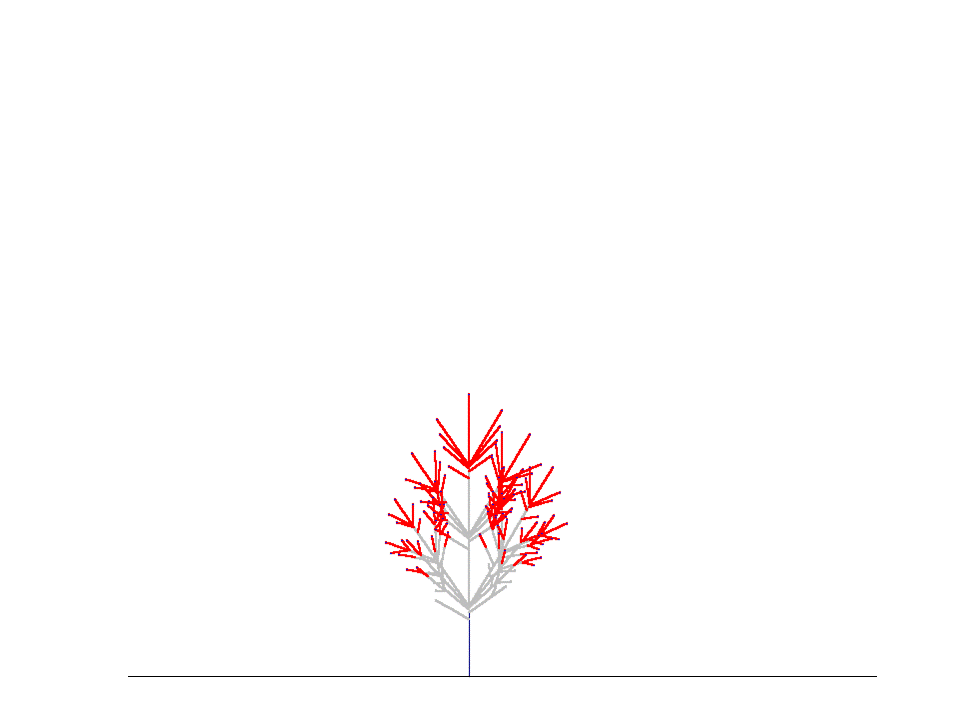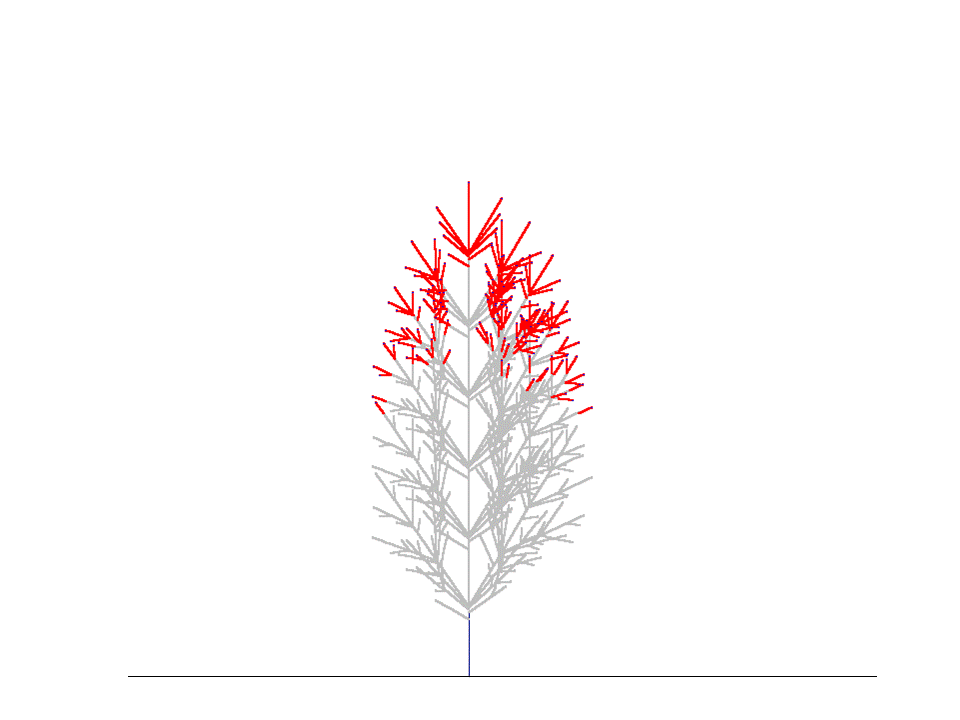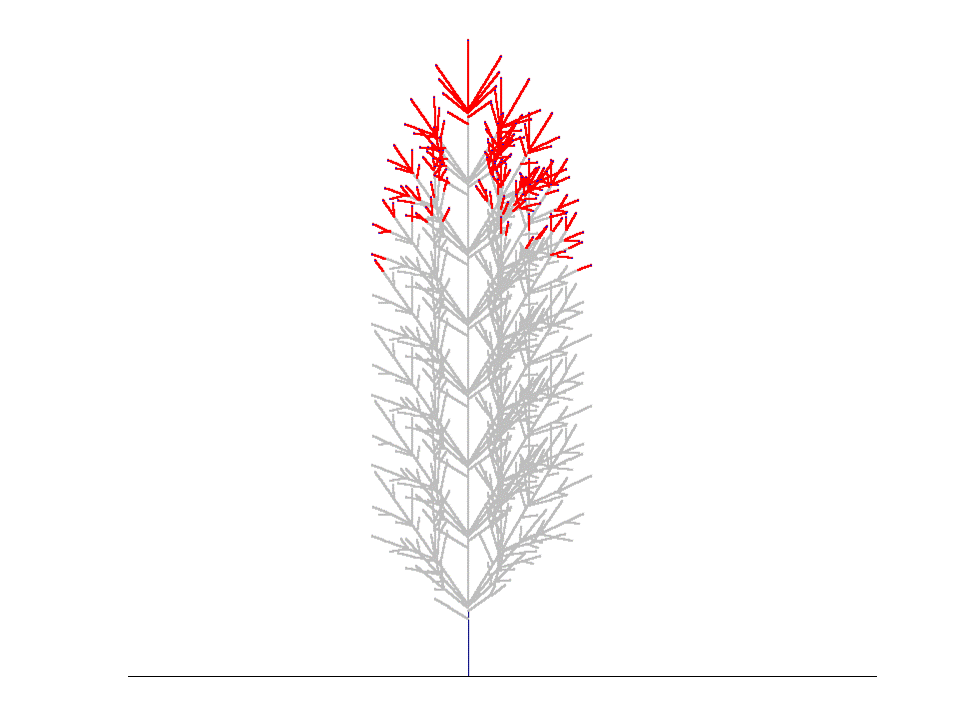
上の結果は、コナラ(Quercus serrata)の小枝を樹冠先端の部分からたくさんとってきて枝分かれ形態のデータをとり、このデータから得られたパラメタを使って枝の分岐構造がどのように発達するかをシミュレートしたときのものです。毎年の枝の状態をみるために、この様子を視覚的に示したのが下の図です。
親枝が発達を初めて4~5年すると、枝先端のシュートが発達を続ける一方で側枝はそのうち死んでしまうため、枝全体としては一定の本数の当年性シュートを保持したまま上向きに発達を続けていく、ということがわかりました。このような、定常的に一定サイズを保った枝のことを「枝葉クラスター "foliage cluster"」と名付けました。(Sumida & Takai (2003), Sumida & Umeki (2007)).
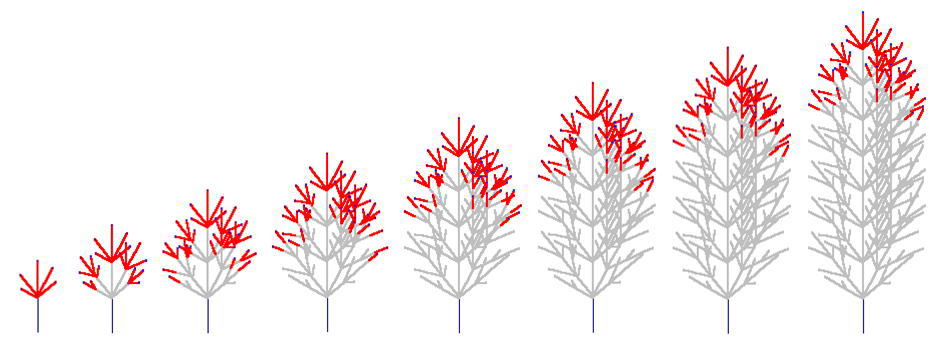
枝葉クラスターをつくって生育を続ける枝のシミュレーション(樹冠最先端のコナラの枝のデータをもとに再現)。枝は左から右へと1年ずつ成長しする。赤い線が当年性シュート(葉は描いてない)。先端に当年性シュートがないグレーの枝は、その枝が死んでいることを意味する。 1年目、5本の当年生(0年生)シュートが親シュートの上に生まれている。5年目になると、1年目に生まれたシュートのうちの下側4本のシュートは死に、親シュートの一番上にあったシュートだけが生き残っている。このとき、枝葉クラスターのサイズは定常状態になり、それ以降は枝葉クラスター内の当年生シュートの数は増えない(下図)。上の図では、5年目に最高齢の4年生シュートの本数は1本だけである。
さきの FIg 1にはトリックがあります。枝葉クラスターが定常状態になった後は、枝葉クラスターの中の最も高齢のシュートの数は1本だけなのです。つまり、Fig 1の、見かけ上「指数関数的に枝分かれ」する構造は、枝葉クラスターの中だけで成立しているのでした。
樹木どうしの競争の実際
もちろん上のような小枝スケールの現象がすべてを説明するわけではありません。
森林の林冠を構成する個体間、種間では競争が起こります。したがって、森林の林冠全体の状態はある程度定常的であっても、林冠を構成する個体は競争によって入れ替わっているかもしれません。ここで言う「競争」とは、葉を広げるための空間をめぐる個体間、個々の枝間の相互作用のことです。.
言うまでもなく、林冠を構成する葉は枝によって空間的に(生理的にも)支えられています。最初に「葉層は時間とともに上に上がっていく」という話をしましたが、その一方で、これまで林冠を構成していた樹木が競争に負けて死んでしまうこともあるわけです。森林全体の林冠構造がどう発達していくかを知るためには、個体の葉を広げる「枝」が群落の中でどのように発達するのかを知っておく必要があります。
岐阜県荘川村の広葉樹林では、岐阜大学の小見山章先生たちのグループの研究から、林冠を構成するクリの木(Castanea crenata)の樹冠の広がりが、まわりの木よりもやや狭い傾向にあるらしいことがでわかっていました。この「狭いらしい」傾向は、個体の樹冠サイズのスケールではあまり明確ではありませんでした(つまり、統計的に有意ではありませんでした)。そこで、個体の中の大枝ごとに解析できるよう、一定の土地区画の中に生育している樹木のすべての枝の分岐構造を三次元的に調べました。
このような、一次枝(各個体の、幹から直接出ている枝のこと)ごとに解析を行うことによりわかったことがあります。クリの一次枝は、その枝に隣接する他の個体の枝との距離(枝の基部間の水平距離)が約5m以内で、しかもその枝がクリ以外の樹種の枝のときにだけ、有意に枝の広がりが狭かったのです。つまり、クリの枝は、他の樹種との「接近戦」に弱いのです。 (Sumida et al., 2002).
なぜでしょうか?その理由まではまだわかっていないのですが、おそらくクリの葉のフェノロジー(春先の葉の展開)が他の樹種よりも遅いことが関係していると考えています。
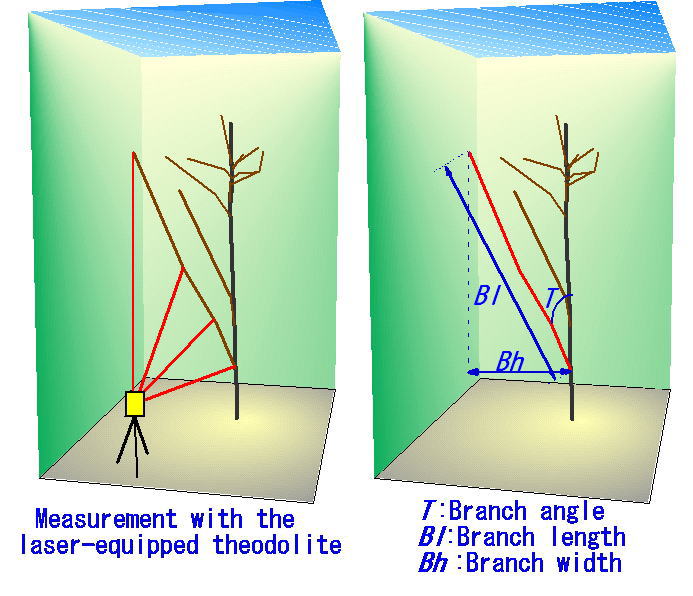

枝の3次元的分枝構造を測定しているところ。(詳しくは Sumida et al (2002))


上の方法で測定し再現した樹木の枝分かれの3D写真:右目に青レンズ、左目に赤レンズでみてね。
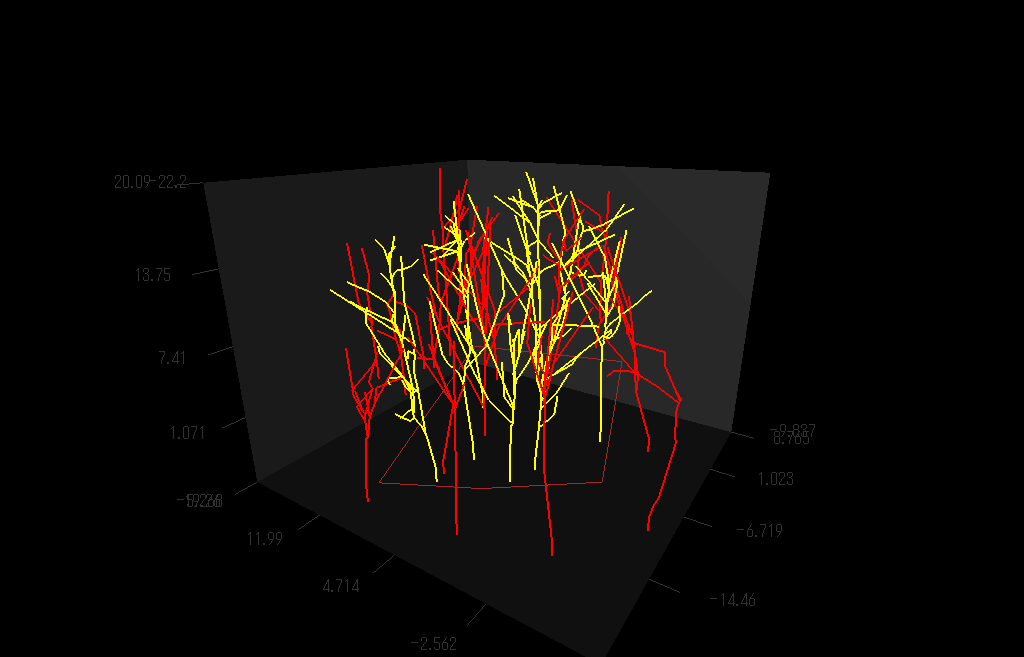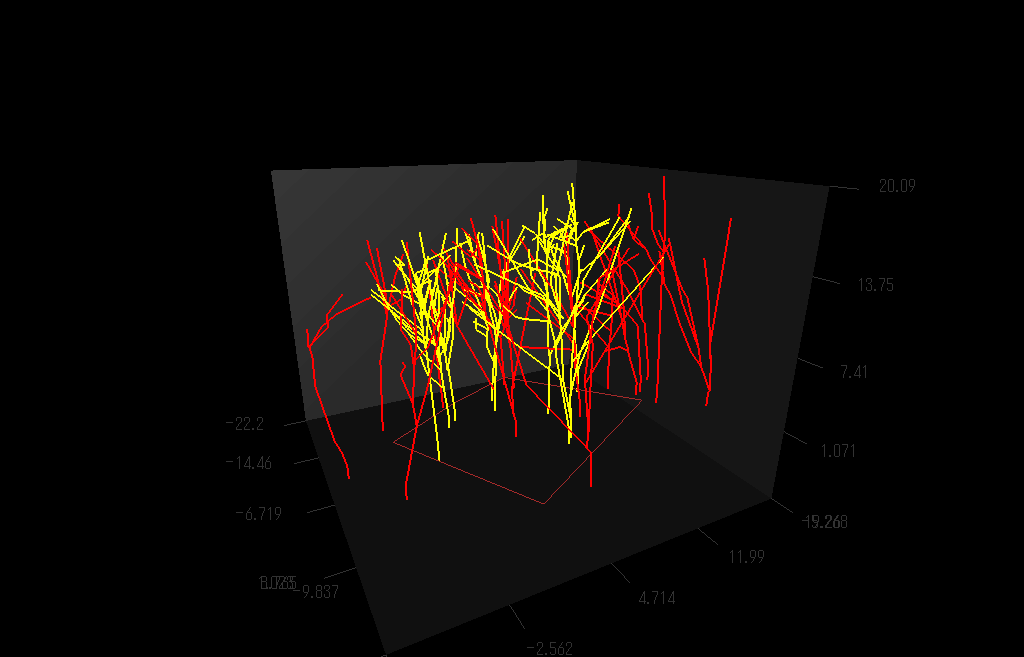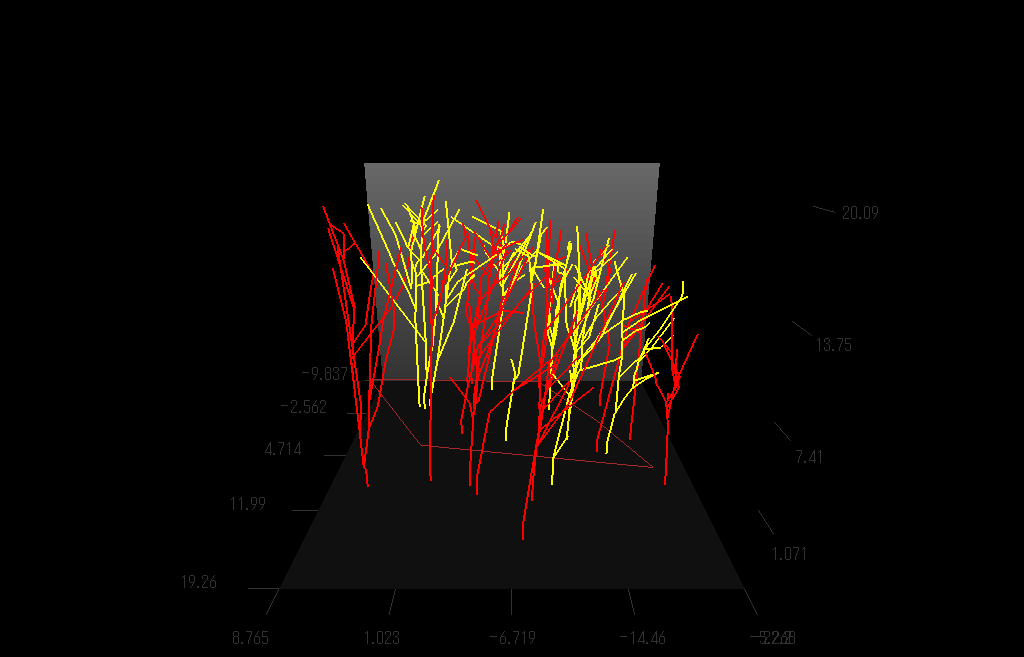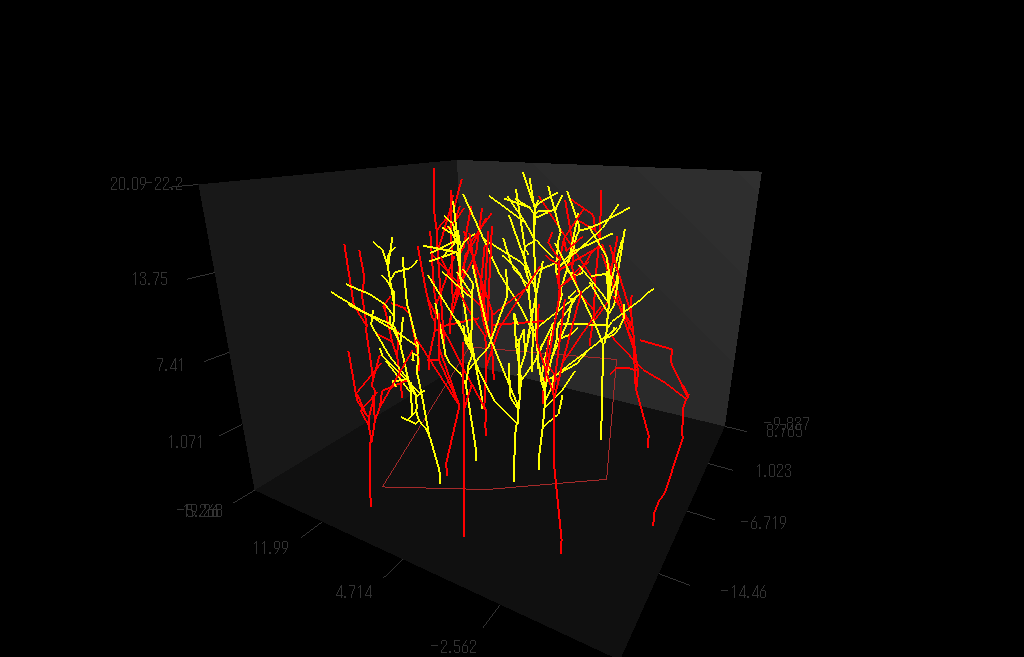
クリ(Castanea crenata)林の幹・枝の構造を再現した図
クリの木(黄色)の枝の広がりは、個々の枝に隣接する枝がクリのものかそれ以外の樹種(赤色)か、によって決まっているようだ。クリ以外の木の枝が隣接している場合には、クリは「接近戦」に弱く、枝の広がりの程度が小さいことがわかった。(Sumida et al. (2002) Annals of Botany 89, 301-310.より)
このような一連の研究のなかで、樹木の小枝スケールの構造や動態から森林全体の構造や動態までのかかわりについて研究をすることが、森林についての理解を深めることになるのだと考えるようになりました。
以上は、私のちょっと前の研究のほんのさわりですが、現在の研究テーマも、この延長上にあります。
森林の構造や動態は,それらを左右する生物的要因(光合成・蒸散など樹木の葉の生理的機能,葉の展開・落葉や幹枝の伸長・太りなどのフェノロジー(生物季節),樹木の個体間競争など)ばかりではなく,樹木や森林を取り巻く物理的環境(光,気温,湿度,雨,風,土壌など)とも深くかかわっています。
より広い視点から樹木・森林について知るため,さまざまな分野の専門家との共同研究を行っています。